| Citation: | Igwilo Christopher Nnaemeka, Egbuna Samuel O, Onoh Maxwell I, Asadu O. Christain, Onyekwulu Chinelo S. Optimization and Kinetic Studies for Enzymatic Hydrolysis and Fermentation of Colocynthis Vulgaris Shrad Seeds Shell for Bioethanol Production[J]. Journal of Bioresources and Bioproducts, 2021, 6(1): 45-64. doi: 10.1016/j.jobab.2021.02.004

|
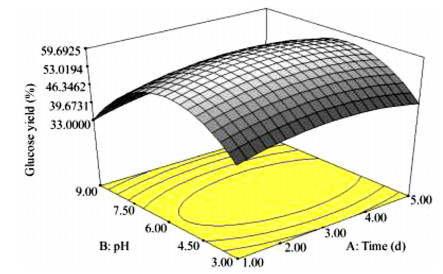
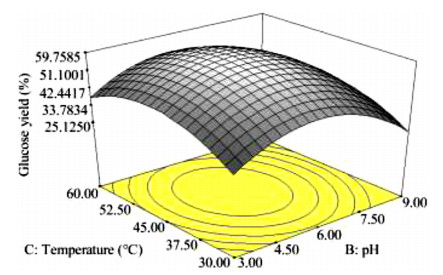
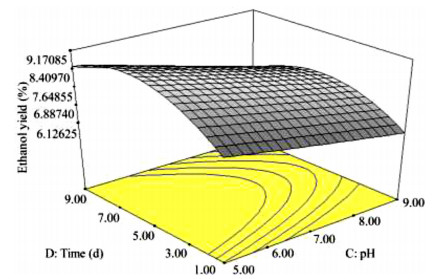
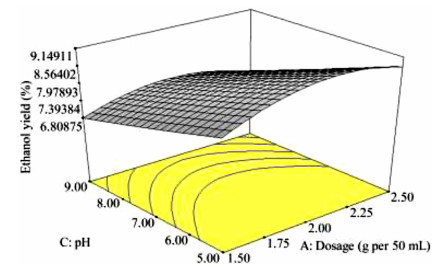
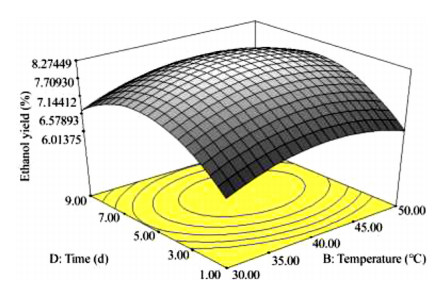
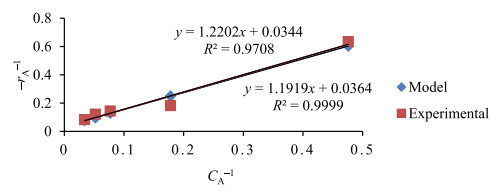
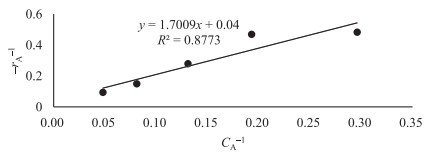
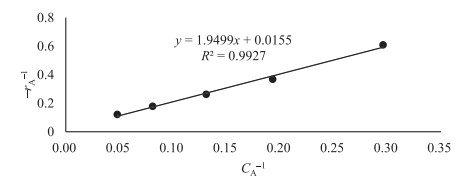
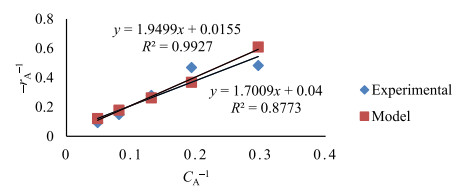
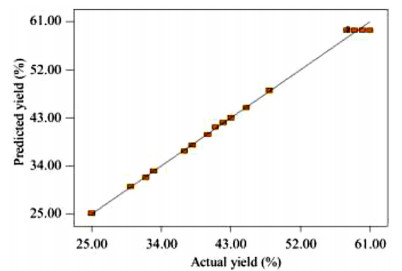
The key process parameters for the hydrolysis and fermentation of Colocynthis vulgaris Shrad seeds shell (CVSSS) were optimized using the Box-Behnken Design (BBD) of Response Surface Methodology (RSM). Kinetic study was also carried out. The proximate analysis of the CVSSS was done by the method of the Association of Organic and Applied Chemistry (AOAC). Enzymatic hydrolysis was experimented by using Aspergillus Niger as a crude enzyme isolated from soil at sawdust dump site and screened for cellulosic activities. Factors that affected the hydrolysis of the CVSSS were screened by using the Greco-Latin square design of experiment. However, for Saccharomyces cerevisiae, factors that affected the fermentation of the CVSSS were screened by using the same Greco-Latin square design of experiment. Meanwhile, the result of the proximate analysis revealed that the CVSSS had 73.54% cellulose which could be converted to bioethanol. It was established that temperature, pH and time had significant effect on hydrolysis, while the optimum results were obtained at 46.8 ℃, 3.32 d, 5.68 and 59.87% for temperature, time, pH and glucose yield, respectively. Temperature, yeast dosage, pH and time had significant effect on fermentation, while the optimum results from optimization were found to be 33.58 ℃, 7.0, 3.55 d, 1.65 g per 50 mL and 25.6% for temperature, pH, time, yeast dosage and ethanol yield, respectively. The kinetics of both the enzymatic hydrolysis and fermentation agreed with the Michealis-Menten kinetic model with the correlation coefficients (R2) of 0.9708 and 0.8773, respectively. However, from the error analysis, the experimental and predicted values had a very good relationship as described by Michaelis-Menten model.
|
Abdulkareem, A. S., Ayo, S. A., Ogochukwu, M. U., 2015. Production and characterization of bioethanol from sugarcane bagasse as alternative energy sources. London U. K. : Proceedings of the World Congress on Engineering.
|
|
Agu, C. M. , Kadurumba, C. H. , Agulanna, A. C. , Aneke, O. O. , Agu, I. E. , Eneh, J. N. , 2018. Nonlinear kinetics, thermodynamics, and parametric studies of Colocynthis vulgaris shrad seeds oil extraction. Ind. Crop. Prod. 123, 386-400. doi: 10.1016/j.indcrop.2018.06.074
|
|
Agu, C. M. , Menkiti, M. C. , Ekwe, E. B. , Agulanna, A. C. , 2020. Modeling and optimization of Terminalia catappa L. kernel oil extraction using response surface methodology and artificial neural network. Artif. Intell. Agric. 4, 1-11. http://www.sciencedirect.com/science/article/pii/S2589721720300064
|
|
Akponah, E. , Akpomie, O. , 2011. Analysis of the suitability of yam, potato and cassava root peels for bioethanol production using saccharomyces cerevisiae. International Research Journal of Microbiology 2, 393-398. http://www.mendeley.com/research/analysis-suitability-yam-potato-cassava-root-peels-bioethanol-production-using-saccharomyces-cerevis/
|
|
Amerine, M. A. , Ough, C. S. , 1974. Wine and Must Analysis. John Wiley & Sons, New York.
|
|
Ana, D. , Julie, L. , Ana, B. , Ignacio, D. , lldefonso, C. , 2013. Pretreatment of rice hulls with alkaline peroxide to enhance enzyme hydrolysis for ethanol production. The Italian Association of Chemical Engineering 32, 23-25. http://www.researchgate.net/publication/273951995_Pretreatment_of_Rice_Hulls_with_Alkaline_Peroxide_to_Enhance_Enzyme_Hydrolysis_for_Ethanol_Production
|
|
Asadu, C. O. , Egbuna, S. O. , Chime, T. O. , Eze, C. N. , Kevin, D. , Mbah, G. O. , Ezema, A. C. , 2019. Survey on solid wastes management by composting: optimization of key process parameters for biofertilizer synthesis from agro wastes using response surface methodology (RSM). Artif. Intell. Agric. 3, 52-61. http://www.sciencedirect.com/science/article/pii/S2589721719300340
|
|
Augustine, O. , Ayeni, O. A. , Adeeyo, O. M. , Oresegun, T. E. O. , 2015. Compositional analysis of lignocellulosic materials: evaluation of an economically viable method suitable for woody and non-woody biomass. American Journal of Engineering Research 4, 14-19. http://www.researchgate.net/publication/280641412_Compositional_analysis_of_lignocellulosic_materials_Evaluation_of_an_economically_viable_method_suitable_for_woody_and_non-woody_biomass
|
|
Carrillo, F. , Lis, M. J. , Colom, X. , Lopez-Mesas, M. , Valldeperas, J. , 2005. Effect of alkali pretreatment on kinetic study of the enzymatic hydrolysis of sugarcane bagasse 447. Brazilian Journal of Chemical Engineering 30, 437-447.
|
|
Chen, G. , Chen, J. , Srinivasakannan, C. , Peng, J. H. , 2012. Application of response surface methodology for optimization of the synthesis of synthetic rutile from titania slag. Appl. Surf. Sci. 258, 3068-3073. doi: 10.1016/j.apsusc.2011.11.039
|
|
Chen, M. , Zhao, J. , Xia, L. M. , 2008. Enzymatic hydrolysis of maize straw polysaccharides for the production of reducing sugars. Carbohydr. Polym. 71, 411-415. doi: 10.1016/j.carbpol.2007.06.011
|
|
Efri, M., Dwi, W. W., Djali, F., Edi, S., 2017. Optimization and kinetic modelling of the enzymatic hydrolysis of oil palm petioles. ICSAFS Conference Proceedings, 2nd International Conference on Sustainable Agriculture and Food Security, A Comprehensive Approach.
|
|
Ezeonu, I. M., Okafor, J. I., Ogbonna, J. C., 2011. Laboratory Exercises in Microbiology. Ephrata Press,
|
|
Farah, A. , Ahmad, T. J. , Mohd, H. K. , Maizirwan, M. , 2011. Study of growth kinetic and modelling of ethanol production by Saccharomyces cerevisae. African Journal of Biotechnology 16, 18842-18846. http://www.cabdirect.org/abstracts/20123030865.html;jsessionid=4AD84BD4D9C0C09C709D1F0BA7A6EEC5
|
|
Fogler, H. S., 2006. Elements of Chemical Reaction Engineering. Prentice Hall of India, New
|
|
Galbe, M. , Zacchi, G. , 2002. A review of the production of ethanol from softwood. Appl. Microbiol. Biotechnol. 59, 618-628. doi: 10.1007/s00253-002-1058-9
|
|
Highina, B. K. , Hashim, I. , Bugaje, I. M. , 2011. Optimization of ethanol production from sugar molasses in Nigeria. Journal of Applied Technology in Environmental Sanitation 1, 233-237. http://www.cabdirect.org/abstracts/20123193149.html
|
|
Horwitz, W., Latima, G., 2005. Official Method of Analysis' appendix C. Association of organic and Applied Chemistry (AOAC) International. Pp 7-12; pp 16-34
|
|
Igbokwe, P. K. , Idogwu, C. N. , Nwabanne, J. T. , 2016. Enzymatic hydrolysis and fermentation of plantain peels: optimization and kinetic studies. Adv. Chem. Eng. Sci. 6, 216-235. doi: 10.4236/aces.2016.62023
|
|
Ighodaro, O. M., 2012. Evaluation Study of Nigerian Species of Musa paradisiaca Peels. Available at: http://www.sciencepub.net/researcher.
|
|
Itelima, J., Onwuliri, F., Onwuliri, E., Onyimba, I., Oforji, S., 2013. Bio-ethanol production from banana, plantain and pineapple peels by simultaneous saccharification and fermentation process. Int. J. Environ. Sci. Dev. 213-216.
|
|
Kitanović, S. , Milenović, D. , Veljković, V. B. , 2008. Empirical kinetic models for the resinoid extraction from aerial parts of St. John's wort (Hypericum perforatum L. ). Biochem. Eng. J. 41, 1-11. doi: 10.1016/j.bej.2008.02.010
|
|
Lalitha, G., Sivaraji, R., 2011. Use of fruit biomass peel for ethanol production. International Journal of Pharma and Bio Science 2, 23.
|
|
Lazic, Z. R., 2004. Design of Experiment in Chemical Engineering. Weinheim: Wiley-VCH Verlag GmbH & Co. KGaA.
|
|
Lebaka, V. R. , Obulam, V. S. , Young, J. W. , 2011. Production of ethanol from mango peel by saccharomyces cerevisiae. Academic Journal 10, 4183-4189. http://www.cabdirect.org/abstracts/20113187787.html
|
|
Luo, X. L., Liu, J., Zheng, P. T., Li, M., Zhou, Y., Huang, L. L., Chen, L. H., Shuai, L., 2019. Promoting enzymatic hydrolysis of lignocellulosic biomass by inexpensive soy protein. Biotechnol. Biofuels 12, 51.
|
|
Mazaheri, H. , Ghaedi, M. , Ahmadi Azqhandi, M. H. , Asfaram, A. , 2017. Application of machine/statistical learning, artificial intelligence and statistical experimental design for the modeling and optimization of methylene blue and Cd(ⅱ) removal from a binary aqueous solution by natural walnut carbon. Phys. Chem. Chem. Phys. 19, 11299-11317. doi: 10.1039/C6CP08437K
|
|
Menkiti, M. C. , Agu, C. M. , Udeigwe, T. K. , 2017. Kinetic and parametric studies for the extractive synthesis of oil from Terminalia catappa L. kernel. React. Kinetics Mech. Catal. 120, 129-147. doi: 10.1007/s11144-016-1101-y
|
|
Mike, B., Sue, A. A., 1998. Fermented fruits and vegetables. A global perspective. Fao Agricultural service Bulletin 134
|
|
Murray R. S., Larry J. S., 2011. Schaum's Outlines Statistics (4th Edition). New York: McGraw Hill.
|
|
NsukkaFarah, A. , Ahmad, T. J. , Mohd, H. K. , Maizirwan, M. , 2011. Study of growth kinetic and modelling of ethanol production by Saccharomyces cerevisae. African Journal of Biotechnology 16, 18842-18846. http://www.cabdirect.org/abstracts/20123030865.html;jsessionid=4AD84BD4D9C0C09C709D1F0BA7A6EEC5
|
|
Ocloo, F. C. K. , Ayernor, G. S. , 2008. Physical, chemical and microbiological changes in alcoholic fermentation of sugar syrup from cassava flour. African Journal of Biotechnology 7, 164-168. http://www.oalib.com/paper/1326307
|
|
Ogbe, A. O., George, G. A. L., 2012. Nutritional and anti-nutrient composition of melon Husks: potential as feed ingredient in poultry Diet
|
|
Ogbonna, O. , 2013. Floral habits and seed production characteristics in Egusi melon (Colocynthis citrullus L. ). J. Plant Breed. Crop Sci. 4, 137-140. doi: 10.5897/JPBCS2013.0381
|
|
Ogbonna, P. E. , Obi, I. U. , 2000. The influence of poultry manure application and plant density on the growth and yield of Egusi melon (Colocynthis citrullus) on the Nsukka Plains of south eastern Nigeria. Agro-Science 1, 63-74. http://www.cabdirect.org/abstracts/20033106736.html
|
|
Ogunwa, K. I. , Ofodile, S. , Achugasim, O. , 2015. Feasibility study of melon seed oil as a source of biodiesel. J. Power Energy Eng. 3, 24-27. http://www.cqvip.com/QK/72737X/20158/HS727372015008003.html
|
|
Onwu, D. O., 2004. Integrated Energy Conversion (1st Edition). Enugu: Immaculate Publications Limited.
|
|
Orjiakor, P. I. , Igborbgor, C. J. , Ogu, G. I. , 2017. Bio-ethanol yielding potentials of melon seed peels using fungal isolates from palm oil effluents. IJBLST 9, 18-25. http://www.researchgate.net/publication/322629139_Bio-ethanol_yielding_potentials_of_Melon_seed_peels_using_fungal_isolates_from_Palm_oil_effluents
|
|
Oyeleke, S. B. , Dauda, B. E. N. , Oyewole, O. A. , Okoliegbe, I. N. , Ojobode. T. , 2012. Production of bioethanol from cassava and sweet potato peels. Advanced in Environmental Biology 6, 241-245. http://www.cabdirect.org/abstracts/20123057510.html
|
|
Pervez, S., Aman, A., Iqbal, S., Siddiqui, N., Ul Qader, S., 2014. Saccharification and liquefaction of cassava starch: an alternative source for the production of bioethanol using amylolytic enzymes by double fermentation process. BMC Biotechnol. 14, 49.
|
|
Pilkington, J. L. , Preston, C. , Gomes, R. L. , 2014. Comparison of response surface methodology (RSM) and artificial neural networks (ANN) towards efficient extraction of artemisinin from Artemisia annua. Ind. Crop. Prod. 58, 15-24. doi: 10.1016/j.indcrop.2014.03.016
|
|
Pornpunyapat, J. , Chotigeat, W. , Chetpattananondh, P. , 2014. Bioethanol production from pineapple peel juice using Saccharomyces cerevisiae. Adv. Mater. Res. 875/876/877, 242-245. http://kns.cnki.net/KCMS/detail/detail.aspx?dbcode=IPFD&filename=XYSW201212001047
|
|
Rosdee, N. M., Masngut, N., Shaarani, S. M., Jamek, S., Sueb, M. M., 2020. Enzymatic hydrolysis of lignocellulosic biomass from pineapple leaves by using endo-1, 4-xylanase: effect of pH, temperature, enzyme loading and reaction time. IOP Conf. Ser. : Mater. Sci. Eng. 736, 022095.
|
|
Sakimoto, K. , Kanna, M. C. , Matsumura, Y. , 2017. Kinetic model of cellulose degradation using simultaneous saccharification and fermentation. Biomass Bioenergy 99, 116-121. doi: 10.1016/j.biombioe.2017.02.016
|
|
Streamer, M. , Eriksson, K. E. , Pettersson, B. , 1975. Extracellular enzyme system utilized by the fungus Sporotrichum pulverulentum (Chrysosporium lignorum) for the breakdown of cellulose. functional characterization of five endo-1, 4-beta-glucanases and one exo-1, 4-beta-glucanase. Eur. J. Biochem. 59, 607-613. doi: 10.1111/j.1432-1033.1975.tb03919.x/full
|
|
Talib, N. B. , Triwahyono, S. , Jalil, A. A. , Mamat, C. R. , Salamun, N. , Fatah, N. A. A. , Sidik, S. M. , Teh, L. P. , 2016. Utilization of a cost effective Lapindo mud catalyst derived from eruption waste for transesterification of waste oils. Energy Convers. Manag. 108, 411-421. doi: 10.1016/j.enconman.2015.11.031
|
|
Tengborg, C. , Galbe, M. , Zacchi, G. , 2001. Influence of enzyme loading and physical parameters on the enzymatic hydrolysis of steam-pretreated softwood. Biotechnol. Prog. 17, 110-117. doi: 10.1021/bp000145+
|
|
Zakpaa, H. D., Mak, M., Johnson, F. S., 2009. Production of bio-ethanol from corncobs using Aspergillus niger and S. cerevisiae in simultaneous saccharification and fermentation. Academic Journal.
|
|
Zivorad, R. L., 2004. Design of Experiment in Chemical Engineering. Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim.
|
Figures(16) / Tables(20)
 WeChat: JournalBandB
WeChat: JournalBandBCopyright © 2019 Editorial Office of Journal of Bioresources and Bioproducts
Supported by: Beijing Renhe Information Technology Co. Ltd support: info@rhhz.net



 DownLoad:
DownLoad: