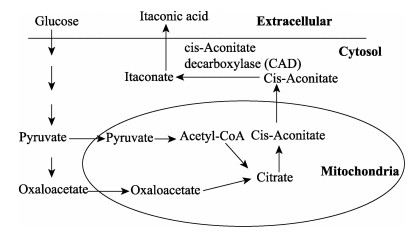
| Citation: | YANG Jing, XU Hao, JIANG Jianchun, ZHANG Ning, XIE Jingcong, WEI Min, ZHAO Jian. Production of Itaconic Acid Through Microbiological Fermentation of Inexpensive Materials[J]. Journal of Bioresources and Bioproducts, 2019, 4(3): 135-142. doi: 10.12162/jbb.v4i3.001

|
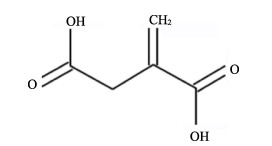
Itaconic acid (IA) is a precursor of various chemicals, which has been on the US Department of Energy's list of the top 12 platform chemicals that can potentially be produced from biomass. The IA is mainly produced through a bio-fermentation method with sugar as raw material. However, the high cost and low yield restrict the industrial application of the IA. This review summarized research advances in the IA production from alternative substrates. In particular, the types of substrates as well as existing problems in the IA fermentation, strain breeding, and process regulation were discussed. And it is pointed that decreasing the price of the IA through use of alternative substrates and increasing the titer of the IA through genetic engineering should be further focused on to support the industrialization of the IA.
|
Bafana R, Pandey R A, 2018. New approaches for itaconic acid production:bottlenecks and possible remedies. Critical Revi-ews in Biotechnology, 38(1):68-82. DOI:10.1080/07388551. 2017.1312268.
|
|
Blazeck J, Hill A, Jamoussi M, et al., 2015. Metabolic engi-neering of Yarrowia lipolytica for itaconic acid production. Metabolic Engineering, 32:66-73. DOI:10.1016/j.ymben. 2015.09.005.
|
|
Blazeck J, Miller J, Pan A, et al., 2014. Metabolic engineering of Saccharomyces cerevisiae for itaconic acid production. Applied Microbiology and Biotechnology, 98(19):8155-8164. DOI: 10.1007/s00253-014-5895-0.
|
|
Choi S, Song C W, Shin J H, et al., 2015. Biorefineries for the production of top building block chemicals and their deri-vatives. Metabolic Engineering, 28:223-239. DOI: 10.1016/j.ymben.2014.12.007.
|
|
Delidovich I, Hausoul P J C, Deng L, et al., 2016. Alternative monomers based on lignocellulose and their use for polymer production. Chemical Reviews, 116(3):1540-1599. DOI: 10.1021/acs.chemrev.5b00354.
|
|
Dowlathabad M, Jaheer M D, Hussain S P V, et al., 2007. Fermentatative production of itaconic acid by Aspergillus terreus using Jatropha seed cake. African Journal of Bio-technology, 6(18):2140-2142. DOI: 10.5897/ajb2007.000-2333.
|
|
EI-Imam A A, Du C Y, 2014. Fermentative itaconic acid pro-duction. Journal of Biodiversity, Bioprospecting and Deve-lopment, 1(2):1-8. http://cn.bing.com/academic/profile?id=ceea78e00fdae9e7bc758de9f3f82643&encoded=0&v=paper_preview&mkt=zh-cn
|
|
EI-Imam A A, Du C Y, 2015. Itaconic acid production from sorghum bran:a biorefining approach. Fundamentals of Renewable Energy and Applications, 5(5):108. DOI: 10.4172/2090-4541.S1.002.
|
|
Geiser E, Przybilla S K, Engel M, et al., 2016. Genetic and biochemical insights into the itaconate pathway of Ustilago maydis enable enhanced production. Metabolic Engineering, 38:427-435. DOI: 10.1016/j.ymben.2016.10.006.
|
|
Gnanasekaran R, Dhandapani B, Gopinath K P, et al., 2018. Synthesis of itaconic acid from agricultural waste using novel Aspergillus niveus. Preparative Biochemistry and Bio-technology, 48(7):605-609. DOI:10.1080/10826068.2018. 1476884.
|
|
Harder B, Bettenbrock K, Klamt S, 2016. Model-based metabolic engineering enables high yield itaconic acid production by Escherichia coli. Metabolic Engineering, 38:29-37. DOI: 10.1016/j.ymben.2016.05.008.
|
|
Hevekerl A, Kuenz A, Vorlop K, 2014. Influence of the pH on the itaconic acid production with Aspergillus terreus. Applied Microbiology and Biotechnology, 98(24):10005-10012. DOI: 10.1007/s00253-014-6047-2.
|
|
Hossain A H, Li A, Brickwedde A, et al., 2016. Rewiring a secondary metabolite pathway towards itaconic acid produ-ction in Aspergillus niger. Microbial Cell Factories, 15:130. DOI: 10.1186/s12934-016-0527-2.
|
|
Jeon H, Cheong D, Han Y, et al., 2016. Itaconic acid production from glycerol using Escherichia coli harboring a random synonymous Codon-substituted 5'-coding region variant of the cadA gene. Biotechnology and Bioengineering, 113(7):1504-1510. DOI: 10.1002/bit.25914.
|
|
Jimenez-Quero A, Pollet E, Zhao M J, et al., 2016. Itaconic and fumaric acid production from biomass hydrolysates by Aspergillus strains. Journal of Microbiology and Biotech-nology, 26(9):1557-1565. DOI: 10.4014/jmb.1603.03073.
|
|
Karaffa L, Díaz R, Papp B, et al., 2015. A deficiency of man-ganese ions in the presence of high sugar concentrations is the critical parameter for achieving high yields of itaconic acid by Aspergillus terreus. Applied Microbiology and Biotechnology, 99(19):7937-7944. DOI: 10.1007/s00253-015-6735-6.
|
|
Kim J, Seo H, Bhatia S K, et al., 2017. Production of itaconate by whole-cell bioconversion of citrate mediated by expression of multiple Cis-aconitate decarboxylase (cadA) genes in Escherichia coli. Scientific Reports, 7:39768. DOI: 10.1038/srep39768.
|
|
Kirimura K, Sato T, Nakanishi N, et al., 1997. Breeding of starch-utilizing and itaconic-acid-producing koji molds by interspecific protoplast fusion between Aspergillus terreus and Aspergillus usamii. Applied Microbiology and Biotechnology, 47(2):127-131. DOI: 10.1007/s002530050900.
|
|
Klement T, Büchs J, 2013. Itaconic acid-A biotechnological process in change. Bioresource Technology, 135:422-431. DOI: 10.1016/j.biortech.2012.11.141.
|
|
Klement T, Milker S, Jager G, et al., 2012. Biomass pretreatment affects Ustilago maydis in producing itaconic acid. Microbial Cell Factories, 11(1):43. DOI: 10.1186/1475-2859-11-43.
|
|
Kobayashi T, 1978. Production of itaconic acid from wood waste. Process Biochemistry, 13(5):15-22. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=1f73d4ff38357820bcbb5e74f1ded431
|
|
Kocabas A, Ogel Z B, Bakir U, 2014. Xylanase and itaconic acid production by Aspergillus terreus NRRL 1960 within a biorefinery concept. Annals of Microbiology, 64(1):75-84. DOI: 10.1007/s13213-013-0634-9.
|
|
Krull S, Eidt L, Hevekerl A, et al., 2017a. Itaconic acid production from wheat chaff by Aspergillus terreus. Process Biochemistry, 63:169-176. DOI:10.1016/j.procbio.2017.08. 010.
|
|
Krull S, Hevekerl A, Kuenz A, et al., 2017b. Process development of itaconic acid production by a natural wild type strain of Aspergillus terreus to reach industrially relevant final titers. Applied Microbiology and Biotechnology, 101(10):4063-4072. DOI: 10.1007/s00253-017-8192-x.
|
|
Kuenz A, Gallenmüller Y, Willke T, et al., 2012. Microbial production of itaconic acid:developing a stable platform for high product concentrations. Applied Microbiology and Biotechnology, 96(5):1209-1216. DOI: 10.1007/s00253-012-4221-y.
|
|
Kuenz A, Krull S, 2018. Biotechnological production of itaconic acid:things you have to know. Applied Microbiology and Biotechnology, 102(9):3901-3914. DOI: 10.1007/s00253-018-8895-7.
|
|
Larsen H, Eimhjellen K, 1955. The mechanism of itaconic acid formation by Aspergillus terreus. 1. The effect of acidity. Biochemical Journal, 60(1):135-139. http://d.old.wanfangdata.com.cn/OAPaper/oai_pubmedcentral.nih.gov_1215665
|
|
Li A, Pfelzer N, Zuijderwijk R, et al., 2013. Reduced by-product formation and modified oxygen availability improve itaconic acid production in Aspergillus niger. Applied Microbiology and Biotechnology, 97(9):3901-3911. DOI: 10.1007/s00253-012-4684-x.
|
|
Li A, van Luijk N, ter Beek M, et al., 2011. A clone-based transcriptomics approach for the identification of genes relevant for itaconic acid production in Aspergillus. Fungal Genetics and Biology, 48(6):602-611. DOI:10.1016/j.fgb. 2011.01.013.
|
|
Li X, Zheng K, Lai C H, et al., 2016. Improved itaconic acid production from undetoxified enzymatic hydrolysate of steam-exploded corn stover using an Aspergillus terreus mutant generated by atmospheric and room temperature plasma. BioResources, 11(4):9047-9058. DOI: 10.15376/biores.11.4.9047-9058.
|
|
Lin Y H, Li Y, Huang M C, et al., 2004. Intracellular expression of Vitreoscilla hemoglobin in Aspergillus terreus to alleviate the effect of a short break in aeration during culture. Bio-technology Letters, 26(13):1067-1072. DOI:10.1023/b:bile. 0000032964.15178.7c.
|
|
Mattey M, 1992. The production of organic acids. Critical Reviews in Biotechnology, 12(1/2):87-132. DOI: 10.3109/07388559209069189.
|
|
Papagianni M, 2007. Advances in citric acid fermentation by Aspergillus niger:biochemical aspects, membrane transport and modeling. Biotechnology Advances, 25(3):244-263. DOI: 10.1016/j.biotechadv.2007.01.002.
|
|
Pedroso G B, Montipó S, Mario D A N, et al., 2017. Building block itaconic acid from left-over biomass. Biomass Con-version and Biorefinery, 7(1):23-35. DOI: 10.1007/s13399-016-0210-1.
|
|
Petruccioli M, Pulci V, Federici F, 1999. Itaconic acid production by Aspergillus terreus on raw starchy materials. Letters in Applied Microbiology, 28(4):309-312. DOI: 10.1046/j.1365-2672.1999.00528.x.
|
|
Reddy C S K, Singh R P, 2002. Enhanced production of itaconic acid from corn starch and market refuse fruits by genetically manipulated Aspergillus terreus SKR10. Bioresource Tech-nology, 85(1):69-71. DOI: 10.1016/S0960-8524(02)00075-5.
|
|
Saha B C, Kennedy G J, 2018. Ninety six well microtiter plate as microbioreactors for production of itaconic acid by six Aspergillus terreus strains. Journal of Microbiological Methods, 144:53-59. DOI: 10.1016/j.mimet.2017.11.002.
|
|
Saha B C, Kennedy G J, Bowman M J, et al., 2019. Factors affecting production of itaconic acid from mixed sugars by Aspergillus terreus. Applied Biochemistry and Biotechnology, 187(2):449-460. DOI: 10.1007/s12010-018-2831-2.
|
|
Saha B C, Kennedy G J, Qureshi N, et al., 2017. Production of itaconic acid from pentose sugars by Aspergillus terreus. Biotechnology Progress, 33(4):1059-1067. DOI: 10.1002/btpr.2485.
|
|
Steen E J, Kang Y S, Bokinsky G, et al., 2010. Microbial production of fatty-acid-derived fuels and chemicals from plant biomass. Nature, 463(7280):559-562. DOI: 10.1038/nature08721.
|
|
Sun T, 2016. Research on the optimize and metabolic flux analysis of itaconic acid by fermentation from wheat bran hydrolysate. Hefei: Hefei University of Technology.
|
|
Tevž G, Benčina M, Legiša M, 2010. Enhancing itaconic acid production by Aspergillus terreus. Applied Microbiology and Biotechnology, 87(5):1657-1664. DOI: 10.1007/s00253-010-2642-z.
|
|
Tippkötter N, Duwe A M, Wiesen S, et al., 2014. Enzymatic hydrolysis of beech wood lignocellulose at high solid contents and its utilization as substrate for the production of biobutanol and dicarboxylic acids. Bioresource Technology, 167:447-455. DOI: 10.1016/j.biortech.2014.06.052.
|
|
Vassilev N, Kautola H, Linko Y, 1992. ImmobilizedAspergillus terreus in itaconic acid production from glucose. Biotech-nology Letters, 14(3):201-206. DOI: 10.1007/bf01023359.
|
|
Weastra S R O, 2013. Determination of market potential for selected platform chemicals: itaconic acid, succinic acid, 2, 5-furandicarboxylic acid. Available at:
|
|
Xu D B, Li W T, 1981. Application of corn starch in fermentation industry. Tianjin Microorganism, 3:59-88. http://d.old.wanfangdata.com.cn/Periodical/spgykj201207050
|
|
Yahiro K, Shibata S, Jia S R, et al., 1997. Efficient itaconic acid production from raw corn starch. Journal of Fermentation and Bioengineering, 84(4):375-377. DOI: 10.1016/s0922-338x(97)89265-3.
|
|
Yang C H, Li P W, 2009. The urgency of research and deve-lopment of itaconic acid application. Shandong Food Fermen-tation, 2:18-20. doi: 10.12162/jbb.v4i3.001
|
|
Yang H, Liang H Q, Liao W, et al., 2003. Breeding of over-producing itaconic acid strains in medium with high concentration of initial sugar by ions implantation. Industrial Microbiology, 33(3):30-32. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=gywsw200303007
|
|
Yang J, 2018. Itaconic acid production from forest residues by Aspergillus terreus. Beijing: Chinese Academy of Forestry.
|
|
Zambanini T, Hosseinpour Tehrani H, Geiser E, et al., 2017. Efficient itaconic acid production from glycerol with Ustilago vetiveriae TZ1. Biotechnology for Biofuels, 10:131. DOI: 10.1186/s13068-017-0809-x.
|
|
Zhang B B, 2018. Assessment of raw material supply capability and energy potential of biomass resources in China. Beijing: China Agricultural University.
|
|
Zhao C, Chen S L, Fang H, 2018. Consolidated bioprocessing of lignocellulosic biomass to itaconic acid by metabolically engineering Neurospora crassa. Applied Microbiology and Biotechnology, 102(22):9577-9584. DOI: 10.1007/s00253-018-9362-1.
|
|
Zhao M L, Lu X Y, Zong H, et al., 2018. Itaconic acid production in microorganisms. Biotechnology Letters, 40(3):455-464. DOI: 10.1007/s10529-017-2500-5.
|
|
Zhu P, 2003. Construction of commercial strain producing itaconic acid from raw starch by protoplast fusion. Nanning: Guangxi University.
|
Figures(3)

Copyright © 2019 Editorial Office of Journal of Bioresources and Bioproducts
Supported by: Beijing Renhe Information Technology Co. Ltd support: info@rhhz.net



 DownLoad:
DownLoad: