| Citation: | CHEN Bingwei, WANG Xinzhou, LENG Weiqi, KAN Yu'na, MEI Changtong, ZHAI Shengcheng. Spectroscopic/Microscopic Elucidation for Chemical Changes During Acid Pretreatment on Arundo donax[J]. Journal of Bioresources and Bioproducts, 2019, 4(3): 192-199. doi: 10.12162/jbb.v4i3.008

|
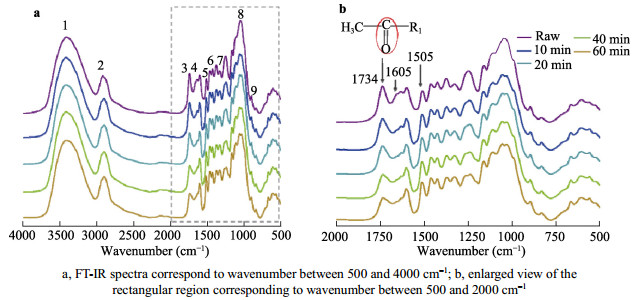
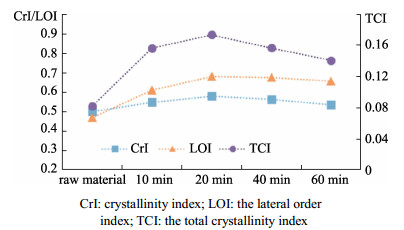
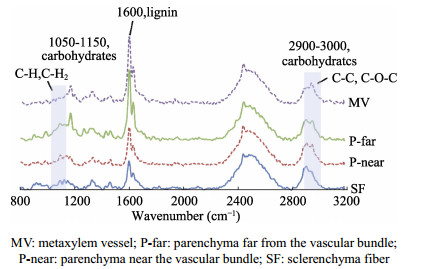
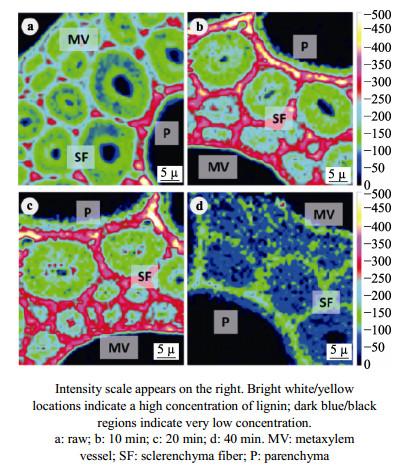
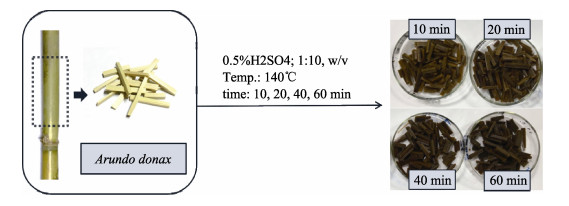
The Arundo donax is a typical fast-growing species from the family Gramineae, which is widely cultivated in China. With a huge yield of A. donax in China, this plant offers great potential for biofuels production. The different types of organization of cell and the tissue in the A. donax could influence the efficiency of enzymatic hydrolysis. In this study, A. donax was subjected to 0.5% (w/w) sulfuric acid (H2SO4) for pretreatment at 140℃ for 10 min, 20 min, 40 min, and 60 min, respectively. The changes in microstructure, chemical composition, topochemical properties were comprehensively analyzed. Using a series of spectroscopic and microscopic techniques including Fourier transform infrared spectroscopy (FT-IR), X-Ray diffraction (XRD), polarized light microscopy (PLM), and confocal Raman microscopy (CRM) to obtain the correlative structural and chemical information. Analysis results of chemical composition, FT-IR spectra and XRD indicated that with increasing reaction time, more hemicellulose and lignin would be removed. Correspondingly, there was an obvious increase of the cellulose relative crystallinity via extending reaction time. Results of the PLM observations showed that the birefringence gradually dimmed due to the diminishing of the cellulose component. Furthermore, the CRM mapping images showed the lignin component in compound middle lamellar (CML) was difficult to remove relatively as compared with that in secondary walls. These results indicated that the combination of spectroscopic and microscopic elucidation could give an insightful understanding of chemical changes in cellular level during pretreatment.
|
Abdul Khalil H P S, Siti Alwani M, Ridzuan R, et al. 2008. Chemical composition, morphological characteristics, and cell wall structure of Malaysian oil palm fibers. Polymer-Plastics Technology and Engineering, 47(3):273-280. DOI: 10.1080/03602550701866840.
|
|
Bergfjord C, Holst B. 2010. A procedure for identifying textile bast fibres using microscopy:Flax, nettle/ramie, hemp and jute. Ultramicroscopy, 110(9):1192-1197. DOI: 10.1016/j.ultramic.2010.04.014.
|
|
Brienzo M, Abud Y, Ferreira S, et al. 2016. Characterization of anatomy, lignin distribution, and response to pretreatments of sugarcane culm node and internode. Industrial Crops and Products, 84: 305-313. DOI: 10.1016/j.indcrop.20youqi-16.01.039.
|
|
Canilha L, Santos V T O, Rocha G J M et al. 2011. A study on the pretreatment of a sugarcane bagasse sample with dilute sulfuric acid. Journal of Industrial Microbiology & Biotechnology, 38(9):1467-1475. DOI: 10.1007/s10295-010-0931-2.
|
|
Castillo R D P, Araya J, Troncoso E, et al. 2015. Fourier transform infrared imaging and microscopy studies of Pinus radiata pulps regarding the simultaneous saccharification and fermentation process. Analytica Chimica Acta, 866:10-20. DOI: 10.1016/j.aca.2015.01.032.
|
|
Chen L J, Li J B, Lu M S, et al. 2016. Integrated chemical and multi-scale structural analyses for the processes of acid pretreatment and enzymatic hydrolysis of corn stover. Carbohydrate Polymers, 141:1-9. DOI:10.1016/j.carbpol. 2015.12.079.
|
|
Chen Y X, Su N, Zhang K T, et al. 2018. Effect of fiber surface treatment on structure, moisture absorption and mechanical properties of Luffa sponge fiber bundles. Industrial Crops and Products, 123:341-352. DOI:10.1016/j.indcrop.2018. 06.079.
|
|
Donohoe B S, Decker S R, Tucker M P, et al. 2008. Visualizing lignin coalescence and migration through maize cell walls following thermochemical pretreatment. Biotechnology and Bioengineering, 101(5):913-925. DOI: 10.1002/bit.21959.
|
|
Engels F. 1998. Alfalfa stem tissues:cell-wall development and lignification. Annals of Botany, 82(5):561-568. DOI: 10.1006/anbo.1998.0705.
|
|
Evert R F. 2006. Esau's plant anatomy: meristems, cells, and tissues of the plant body: their structure, function, and development (3rd edition). Hoboken, New Jersey: John Wiley & Sons, Inc.
|
|
Gierlinger N, Keplinger T, Harrington M. 2012. Imaging of plant cell walls by confocal Raman microscopy. Nature Protocols, 7(9):1694-1708. DOI: 10.1038/nprot.2012.092.
|
|
Gu H Q, An R X, Bao J. 2018. Pretreatment refining leads to constant particle size distribution of lignocellulose biomass in enzymatic hydrolysis. Chemical Engineering Journal, 352:198-205. DOI: 10.1016/j.cej.2018.06.145.
|
|
Haugan E, Holst B. 2013. Determining the fibrillar orientation of bast fibres with polarized light microscopy:the modified Herzog test (red plate test) explained. Journal of Microscopy, 252(2):159-168. DOI: 10.1111/jmi.12079.
|
|
Himmel M E, Ding S Y, Johnson D K, et al. 2007. Biomass recalcitrance:engineering plants and enzymes for biofuels production. Science, 315(5813):804-807. DOI: 10.1126/science.1137016.
|
|
Horikawa Y, Hirano S, Mihashi A, et al. 2019. Prediction of lignin contents from infrared spectroscopy:chemical digestion and lignin/biomass ratios of Cryptomeria japonica. Applied Biochemistry and Biotechnology. DOI: 10.1007/s12010-019-02965-8
|
|
Ibbett R, Gaddipati S, Davies S, et al. 2011. The mechanisms of hydrothermal deconstruction of lignocellulose:New insights from thermal-analytical and complementary studies. Bioresource Technology, 102(19):9272-9278. DOI:10.1016/j.biortech. 2011.06.044.
|
|
Ji Z, Ma J F, Xu F. 2014. Multi-scale visualization of dynamic changes in poplar cell walls during alkali pretreatment. Microscopy and Microanalysis, 20(2):566-576. DOI: 10.1017/s1431927614000063.
|
|
Ji Z, Zhang X, Ling Z, et al. 2015. Visualization of Miscanthus ×giganteus cell wall deconstruction subjected to dilute acid pretreatment for enhanced enzymatic digestibility. Biotechnology for Biofuels, 8:103. DOI: 10.1186/s13068-015-0282-3.
|
|
Ji Z, Zhang X, Ling Z, et al. 2016. Tissue specific response of Miscanthus ×giganteus to dilute acid pretreatment for enhancing cellulose digestibility. Carbohydrate Polymers, 154:247-256. DOI: 10.1016/j.carbpol.2016.06.086.
|
|
Karimi K, Taherzadeh M J. 2016. A critical review of analytical methods in pretreatment of lignocelluloses:Composition, imaging, and crystallinity. Bioresource Technology, 200:1008-1018. DOI: 10.1016/j.biortech.2015.11.022.
|
|
Ling Z, Chen S, Zhang X, et al. 2017. Exploring crystalline-structural variations of cellulose during alkaline pretreatment for enhanced enzymatic hydrolysis. Bioresource Technology, 224:611-617. DOI: 10.1016/j.biortech.2016.10.064.
|
|
Loow Y L, Wu T Y, Md Jahim J, et al. 2016. Typical conversion of lignocellulosic biomass into reducing sugars using dilute acid hydrolysis and alkaline pretreatment. Cellulose, 23(3):1491-1520. DOI: 10.1007/s10570-016-0936-8.
|
|
Mou H Y, Li B, Fardim P. 2014. Pretreatment of corn stover with the modified hydrotropic method to enhance enzymatic hydrolysis. Energy & Fuels, 28(7):4288-4293. DOI: 10.1021/ef5001634.
|
|
Mou H Y, Orblin E, Kruus K, et al. 2013. Topochemical pretreatment of wood biomass to enhance enzymatic hydrolysis of polysaccharides to sugars. Bioresource Technology, 142:540-545. DOI: 10.1016/j.biortech.2013.05.046.
|
|
Murata Y, Kubo S, Togawa E, et al. 2015. Detection of vascular bundles using cell wall birefringence on exposure to polarized light. Industrial Crops and Products, 65:190-197. DOI: 10.1016/j.indcrop.2014.11.055.
|
|
Oh S Y, Yoo D I, Shin Y, et al. 2005. FTIR analysis of cellulose treated with sodium hydroxide and carbon dioxide. Carbohydrate Research, 340(3):417-428. DOI:10.1016/j. carres.2004.11.027.
|
|
Phitsuwan P, Sakka K, Ratanakhanokchai K. 2016. Structural changes and enzymatic response of Napier grass (Pennisetum purpureum) stem induced by alkaline pretreatment. Bioresource Technology, 218:247-256. DOI:10.1016/j.biortech.2016.06. 089.
|
|
Raj T, Gaur R, Lamba B Y, et al. 2018. Characterization of ionic liquid pretreated plant cell wall for improved enzymatic digestibility. Bioresource Technology, 249:139-145. DOI: 10.1016/j.biortech.2017.09.202.
|
|
Richter S, Müssig J, Gierlinger N. 2011. Functional plant cell wall design revealed by the Raman imaging approach. Planta, 233(4):763-772. DOI: 10.1007/s00425-010-1338-z.
|
|
Simonović Radosavljević J, Bogdanović Pristov J, Lj Mitrović A, et al. 2017. Parenchyma cell wall structure in twining stem of Dioscorea balcanica. Cellulose, 24(11):4653-4669. DOI: 10.1007/s10570-017-1460-1.
|
|
Singh J, Suhag M, Dhaka A. 2015. Augmented digestion of lignocellulose by steam explosion, acid and alkaline pretreatment methods:A review. Carbohydrate Polymers, 117:624-631. DOI: 10.1016/j.carbpol.2014.10.012.
|
|
Sluiter A, Hames B, Ruiz R, et al. 2012. Determination of structural carbohydrates and lignin in biomass. US:National Renewable Energy Laboratory, 1-15. http://cn.bing.com/academic/profile?id=87e2a1ead9896f3d18546ac3742babbb&encoded=0&v=paper_preview&mkt=zh-cn
|
|
Sun Q N, Foston M, Sawada D, et al. 2014. Comparison of changes in cellulose ultrastructure during different pretreatments of poplar. Cellulose, 21(4):2419-2431. DOI: 10.1007/s10570-014-0303-6.
|
|
Thygesen L G, Hidayat B J, Johansen K S, et al. 2011. Role of supramolecular cellulose structures in enzymatic hydrolysis of plant cell walls. Journal of Industrial Microbiology & Biotechnology, 38(8):975-983. DOI: 10.1007/s10295-010-0870-y.
|
|
Tjeerdsma B F, Militz H. 2005. Chemical changes in hydrothermal treated wood:FTIR analysis of combined hydrothermal and dry heat-treated wood. Holz Als Roh-Und Werkstoff, 63(2):102-111. DOI: 10.1007/s00107-004-0532-8.
|
|
Wu X X, Huang C, Zhai S C, et al. 2018. Improving enzymatic hydrolysis efficiency of wheat straw through sequential autohydrolysis and alkaline post-extraction. Bioresource Technology, 251:374-380. DOI:10.1016/j.biortech.2017.12. 066.
|
|
Yang G, Wang J L. 2019. Ultrasound combined with dilute acid pretreatment of grass for improvement of fermentative hydrogen production. Bioresource Technology, 275:10-18. DOI: 10.1016/j.biortech.2018.12.013.
|
|
You T T, Zhang L M, Xu F. 2016. Progressive deconstruction of Arundo donax Linn. to fermentable sugars by acid catalyzed ionic liquid pretreatment. Bioresource Technology, 199:271-274. DOI: 10.1016/j.biortech.2015.08.152.
|
|
Yu Z, Li J L, Li G Y. 2009. Analysis on the potential capacity of exploiting giant reed as an energy forage. Pratacultural Science, 26(6):62-69. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=caoyekx200906012
|
|
Zhai S C, Horikawa Y, Imai T, et al. 2014. Cell wall ultrastructure of palm leaf fibers. IAWA Journal, 35(2):127-137. DOI: 10.1163/22941932-00000054.
|
|
Zhang X M, Qu T J, Mosier N S, et al. 2018. Cellulose modification by recyclable swelling solvents. Biotechnology for Biofuels, 11:191. DOI: 10.1186/s13068-018-1191-z.
|
|
Zhu L, O'Dwyer J P, Chang V S, et al. 2008. Structural features affecting biomass enzymatic digestibility. Bioresource Technology, 99(9):3817-3828. DOI:10.1016/j.biortech. 2007.07.033.
|
Figures(6) / Tables(2)

Copyright © 2019 Editorial Office of Journal of Bioresources and Bioproducts
Supported by: Beijing Renhe Information Technology Co. Ltd support: info@rhhz.net



 DownLoad:
DownLoad: